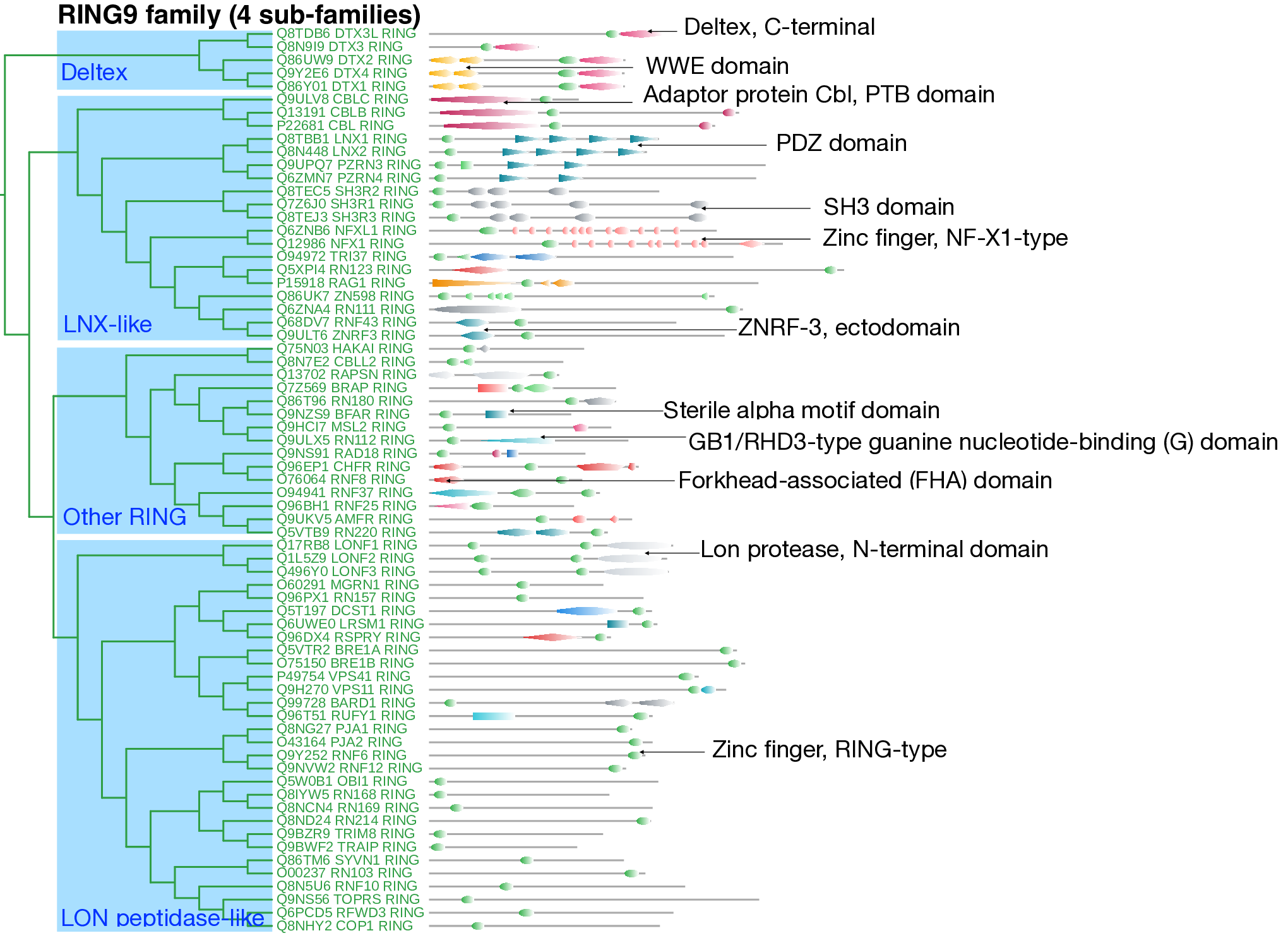
The RING9 family comprises 69 E3 ligases with sparse and diverse InterPro domain annotations. The Zn-finger RING-type domain is often the primary annotated feature within this family. Based on domain architecture and sequence similarity, the family is further categorized into four distinct subfamilies.
The first subfamily consists of five Deltex family members, characterized by the presence of the Deltex C-terminal domain (IPR039396), which contains an ADP-ribose-binding pocket and plays a key role in mediating cell-to-cell communication. Additionally, these proteins possess a WWE domain that recognizes poly-ADP-ribosylated substrates and plays a crucial role in DNA repair. These domain-associated functionalities align with the observed enrichment in biological processes such as the Notch signaling pathway and the cellular component associated with sites of DNA damage.
The remaining three subfamilies exhibit diverse domain architectures. The subfamily termed "LNX-like" includes ligases such as CBL, Ligand of Numb protein-X (LNX), SH3RF, and NFX. Within this group, CBL ligases are distinguished by an N-terminal Cbl adaptor protein with a PTB domain (IPR024159). Members of the LNX family are characterized by PDZ domain repeats (IPR001478), which are typically found in signaling proteins localized to highly specialized submembranous sites. SH3 domain-containing ligases are implicated in apoptosis, whereas NFX ligases contain NF-X1-type repeats, highlighting diverse functional roles within this subfamily.
The "Other RING" subfamily includes ligases that contain GB1/RHD3-type guanine nucleotide (G) binding domains (IPR030386), found in RN112 ligases, as well as N-terminal FHA domains, observed in CHFR and RNF8 ligases. Additionally, this subfamily includes ligases such as LRSM1 and BFAR, which feature a sterile alpha motif (SAM) domain.
The "LON peptidase-like" subfamily includes E3 ligases such as LON peptidase, BRE, and VPS. LON peptidase ligases are notable for containing two RING-type Zn-finger domains and an N-terminal Lon protease domain, which is involved in the degradation of short-lived regulatory and damaged proteins. Other ligases in this group contain only the Zn-finger RING domain.
This heterogeneity in sequence and domain composition is reflected in the multiple sequence alignment (MSA) of the family, which exhibits numerous gaps. The RING9 family is enriched in biological processes such as protein K6-linked ubiquitination, positive regulation of catabolic processes, and DNA metabolic processes. Additionally, RING9 ligases are predominantly localized to the PML body, the site of DNA damage, and the ER quality control compartments.
These ligases are also associated with molecular functions such as histone ubiquitin ligase activity and SH3 domain binding. The PDB contains 144 structures covering 32 out of 69 RING9 proteins. Most RING9 E3 ligases are likely to function as standalone ligases, with only five operating as part of a complex, while the remaining E3s remain unclassified.
Analysis of the ESI network for this family revealed 296 unique E3-specific substrates and 66 family-specific substrates. Small molecule binders were identified specific to CBLB, which form 3 neat segregated clusters, indicating that a similar scaffold interacting with presumably the same binding site can be optimized for handle design.
For updated information and bibliography, please consult the supplementary information in Dutta et al. (2025, preprint)
The values represent E3 ligase confidence scores, calculated as the fraction of databases that annotate the protein as an E3 ligase (see preprint).