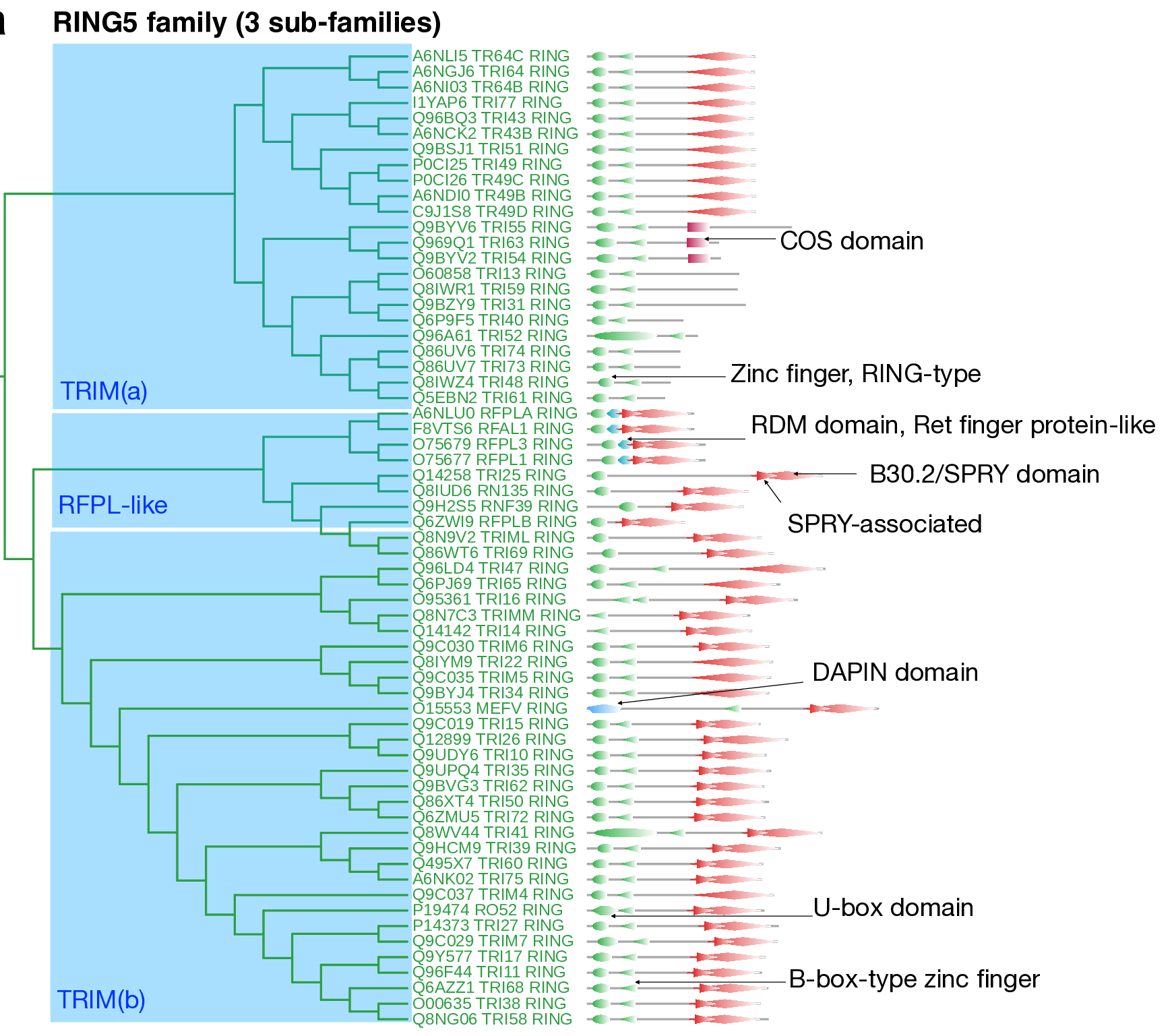
The RING5 family is a highly homogeneous family containing 63 E3s. Most RING5 members are TRIM E3 ligases. Three subfamilies were observed in this family, two for TRIM ligases and one for RFLP ligases. In this family, the catalytic Zn-finger RING-type or Zn-finger B-box-type (IPR000315) domains are typically found near the N-terminus. More than 80% of the RING5 family feature a B30.2/SPRY domain (IPR001870), a known protein–protein interaction domain with three highly conserved motifs (–LDP–, –LDYE–, and –WEVE–). Flanking the SPRY domain at the N-terminus is the SPRY-associated domain (IPR006574). This unique domain architecture is a basis for categorizing TRIM ligases into two distinct subfamilies.
The structural organization of the TRIM ligase in the RING5 family reveals a clear dichotomy: one subfamily designated TRIM(a) features TRIM ligases with a RING-type domain, B-box motif, and B30.2/SPRY domain. These proteins are characterized by a COS domain but notably lack the C-terminal B30.2/SPRY domain. Conversely, TRIM(b) ligases are distinguished by the presence of both SPRY-associated and B30.2/SPRY domains at their C-terminus. The protein Pyrin (MEFV) from the TRIM(b) subfamily shows the presence of a distinct DAPIN domain at the N-terminal.
The DAPIN domain is hypothesized to function as an adaptor involved in the coupling between apoptosis and other immune system disorder pathways.
The other subfamily, "RFPL-like", contains the RFPL protein and a few TRIM ligases. They show the presence of both the SPRY-associated domain, B30.2/SPRY domain, and the RING domains at the C-terminus. However, they contain an RDM domain in between.
The RING5 family is highly homogenous for TRIM family proteins and the SPRY domains. The sequence alignment shows the conserved catalytic RING domain, B-box domains, SPRY associated, and B30.2 SPRY domain. TRIMs are defined by their multimodular structure, including a conserved RING–B-box–coiled-coil (RBCC) domain and variable C-terminal regions. Despite sharing a similar domain architecture, they perform various functions and are involved in antiviral defense and cancer.
The RING5 family demonstrates significant functional segregation. It shows enrichment in the biological process, such as innate immune response, suppression of viral release by the host, and regulation of canonical NF-κB signal transduction. Their most enriched cellular component is the cytoplasmic stress granule, and they are involved in molecular functions such as transcription coregulator activity and ubiquitin-protein ligase activity.
The PDB contains 61 structures covering 21 out of 63 RING5 proteins. Most RING5 family proteins are likely to function as standalone ligases; only eight are presumed to function as complex, and seven E3s are unclassified. Analysis of the ESI network for this family revealed 115 unique E3-specific substrates and 21 family-specific substrates. No existing PROTACs or E3 binders are identified for RING5 family proteins.
For updated information and bibliography, please consult the supplementary information in Dutta et al. (2025, preprint)
The values represent E3 ligase confidence scores, calculated as the fraction of databases that annotate the protein as an E3 ligase (see preprint).