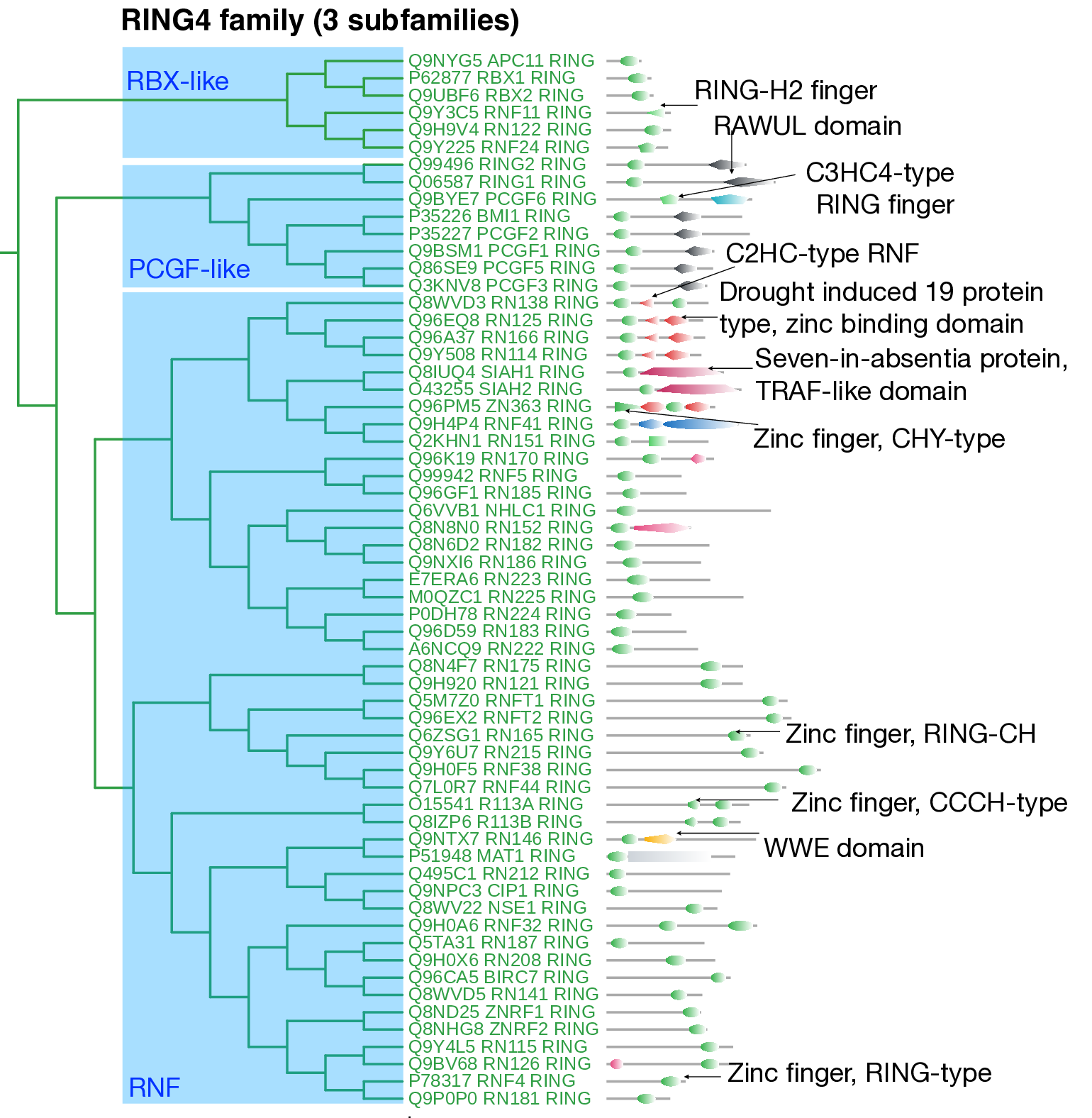
The RING4 family consists of 61 proteins, with more than 50% classified as RING finger proteins (RNF) containing the Zn-finger, RING-type catalytic domain. A notable feature of this family is the clustering of ligases based on the position of their catalytic domains, with those containing C-terminal and N-terminal catalytic domains forming distinct subfamilies. This clustering pattern reflects sequence-level similarity. In total, three subfamilies were identified within this family.
The first subfamily comprises small E3 ligases, including RBX proteins (CRLs) and RNF ligases. The second subfamily consists of eight ligases, corresponding to the Polycomb group (PcG) protein family (PCGF).
The "PCGF-like" subfamily is characterized by the presence of a C-terminal RAWUL domain (IPR032443), which mediates interactions with Cbx members of the PRC1 (Polycomb Repression Complex 1) and facilitates homodimer formation.
The third subfamily contains 47 RNF ligases with diverse domain architectures, including C2HC-type RNF Zn-finger domains, drought-induced 19 protein-type zinc-binding domains, and Zn-finger SIAH-type profiles (IPR013010), and it shows further internal subdivisions. The C-terminal region of SIAH ligases is thought to function as a substrate-and-cofactor-interaction domain (substrate-binding domain, or SBD), facilitating interactions with various proteins. Twelve RNF proteins containing HC/C3H2C3-type RING domains form a distinct subgroup within this subfamily. Some proteins (RN182, RN183, RN186, RN152) also contain transmembrane domains.
The remaining RNF proteins are further categorized based on the location of their RING domains: the N-terminal RING domain is observed in MAT1, RN146, and RN212, while the C-terminal RING domain is present in BIRC7, RN41, and RN126. Additionally, proteins such as RN175, RN121, and RNFTs contain transmembrane domains, likely contributing to their localization within the endoplasmic reticulum (ER).
The RING4 family exhibits a well-conserved RING-type domain, as demonstrated in the multiple sequence alignment (MSA), often occurring in combination with diverse non-catalytic domains. Functionally, this family demonstrates significant segregation. Enrichment analysis indicates involvement in biological processes such as the positive regulation of response to ER stress, protein K11-linked ubiquitination, and the ER-associated degradation (ERAD) pathway. Members of this family are enriched in the PRC1 complex and the ubiquitin ligase complex. Consistent with this, their enriched molecular functions include cullin family protein binding (CRLs), histone ubiquitin ligase activity, and promoter-specific chromatin binding.
The PDB contains 149 resolved structures covering 27 out of 61 RING4 proteins. Most RING4 family proteins are predicted to function as standalone ligases. In contrast, only 13 proteins—primarily from the RBX-like and PCGF-like subfamilies—are expected to function as multi-subunit complexes.
Analysis of the ESI network for this family revealed 215 unique E3-specific substrates and 34 family-specific substrates. Among the small molecules targeting this family of proteins, we identified existing PROTACs and E3 handles for RNF4 and RN114, which could be repurposed to target the co-clustered proteins RN181 and RN166, respectively. Similarly, the new E3 binders identified for proteins MAT1, BMI1, and BIRC7 can be repurposed or developed into potential E3 handles for RN146, PCGF2, and RN141.
For updated information and bibliography, please consult the supplementary information in Dutta et al. (2025, preprint)
The values represent E3 ligase confidence scores, calculated as the fraction of databases that annotate the protein as an E3 ligase (see preprint).