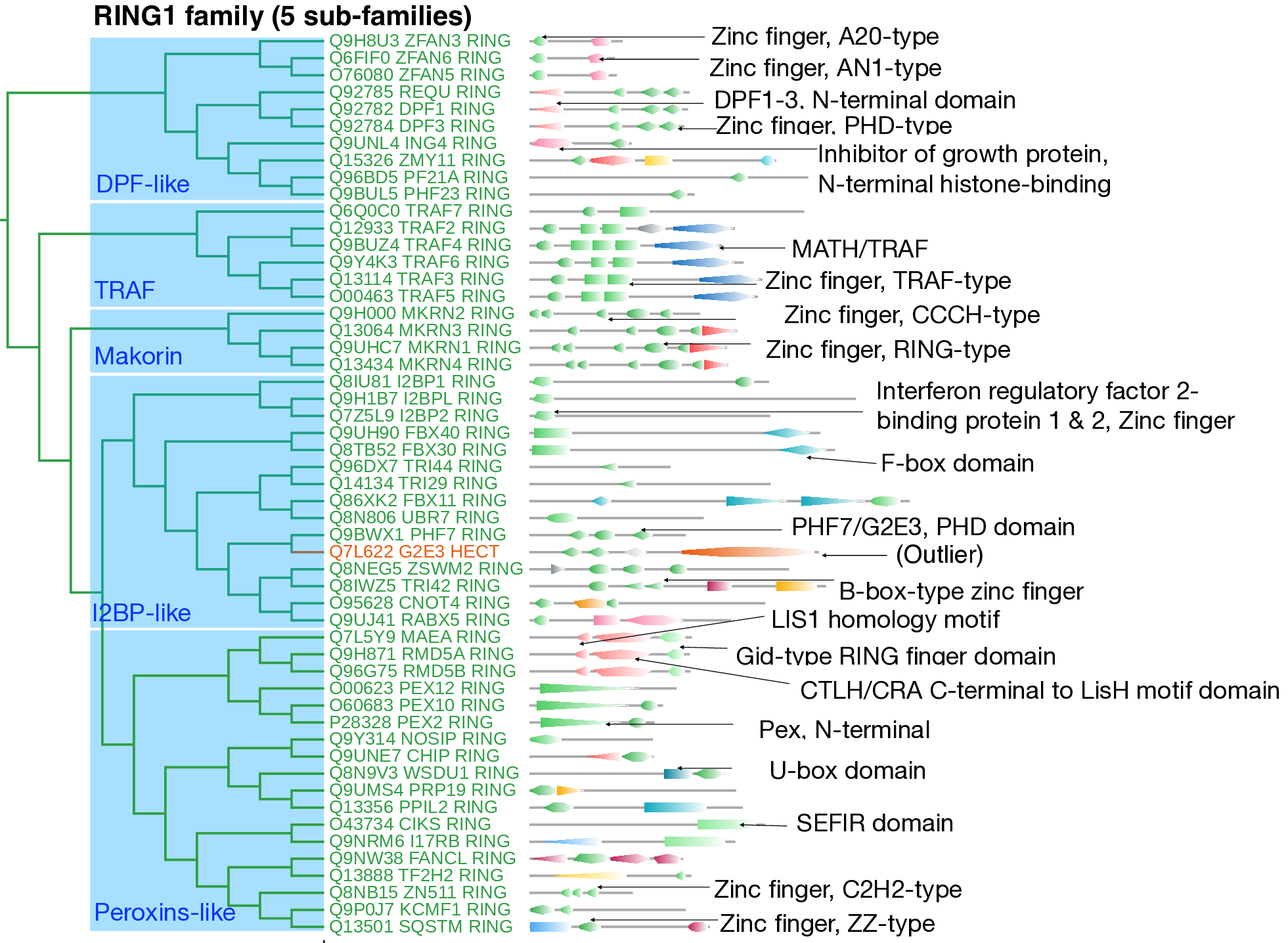
The RING1 family encompasses 53 RING E3 ligases characterized by one or more catalytic Zinc-finger (Zn-finger) domains, including RING-type (IPR001841), PHD-type (IPR001965), or TRAF-type (IPR001293) domains. This family is organized into five distinct subfamilies.
The first subfamily comprises six TNF receptor-associated E3s (TRAFs) that share a unique domain architecture consisting of TRAF-type Zn-finger domains followed by MATH domains. These TRAF ligases are functionally associated with MAPK and JNK signaling pathways, innate immunity, and apoptosis.
The second subfamily consists of four Makorin proteins (MKRN1–4), which are putative ribonucleoproteins characterized by a conserved RING-type Zn-finger domain.
The remaining three subfamilies, containing diverse E3s with varied domain architectures, are named after their well-known members. The family-level multiple-sequence alignment reveals significant heterogeneity within the DPF-like, I2BP-like, and Peroxin-like subfamilies, as evidenced by numerous gaps.
The DPF-like subfamily includes proteins characterized by the DPF domain (double PHD finger proteins, DPF1, DPF3) and members containing tandem repeats of AN20-type and AN1-type Zn-finger domains (ZFAN3, ZFAN5, ZFAN6).
The I2BP-like subfamily encompasses proteins from the interferon regulatory factor 2 binding protein (I2BP) family, F-box proteins (FBX30, FBX40, FBX11), and tripartite motif (TRIM) proteins (TRI44 and TRI29), along with UBR7, ZSWM2, RABX5, and the outlier G2E3. Despite its characteristic C-terminal HECT domain, G2E3's presence in this subfamily is explained by its preserved N-terminal domains: an ePHD domain (IPR042012), Ring-type Zn-finger domain, and PHD-PHF7-G2-H3-like (IPR042012) domain, which are also found in the RING E3 ligase PHF7.
The Peroxins-like subfamily exhibits diverse domain architectures. Three E3s (PEX2, PEX10, PEX12) with well-preserved Peroxin N-terminal domains are involved in peroxisomal protein import. RMD5A, RMD5B, and MAEA proteins form a distinct sub-group characterized by GID-type Zn-finger and LIS1 homology motifs. The remaining members include the nitric oxide synthase-interacting protein (NOSIP), Cys2His2 Zn-finger proteins (CHIP), and other E3s (CIKS, I17RB, FANCL) containing various Zn-finger domain types (C3HC4-, C2H2-, and RING-type).
The RING1 family demonstrates significant functional segregation, with enrichment in specific biological processes, including regulation of canonical NF-κB signal transduction, K63-linked ubiquitination, and protein localization to peroxisomes. At the cellular component level, the family shows enrichment in various complexes, including the CD40 receptor complex, GID complex, nBAF complex, and peroxisomal membrane. The family is also associated with molecular functions such as thioesterase binding, ubiquitin-ubiquitin ligase activity, kinase binding, and protein transmembrane transporter activity.
The PDB contains approximately 168 structures covering 27 of the 53 RING1 proteins. Within this family, sixteen E3s function as complex E3s, including FBX11, FBX30, and FBX40, which are known to be part of E3 complexes that ubiquitinate BCL6, DTL, and other substrates.
Seventeen ligases operate as standalone E3 ligases, and the remaining E3s are currently unclassified. Analysis of the ESI network for this family revealed 191 unique E3-specific substrates and 19 family-specific substrates. We identified seven small molecule binders for ZZ-type Zn-finger-containing protein, SQSTM (p63), with the potential for the rational design of E3 handles and PROTACs. These molecules could also target other co-clustered E3 within the RING1 family, such as KCMF1.
For updated information and bibliography, please consult the supplementary information in Dutta et al. (2025, preprint)
The values represent E3 ligase confidence scores, calculated as the fraction of databases that annotate the protein as an E3 ligase (see preprint).