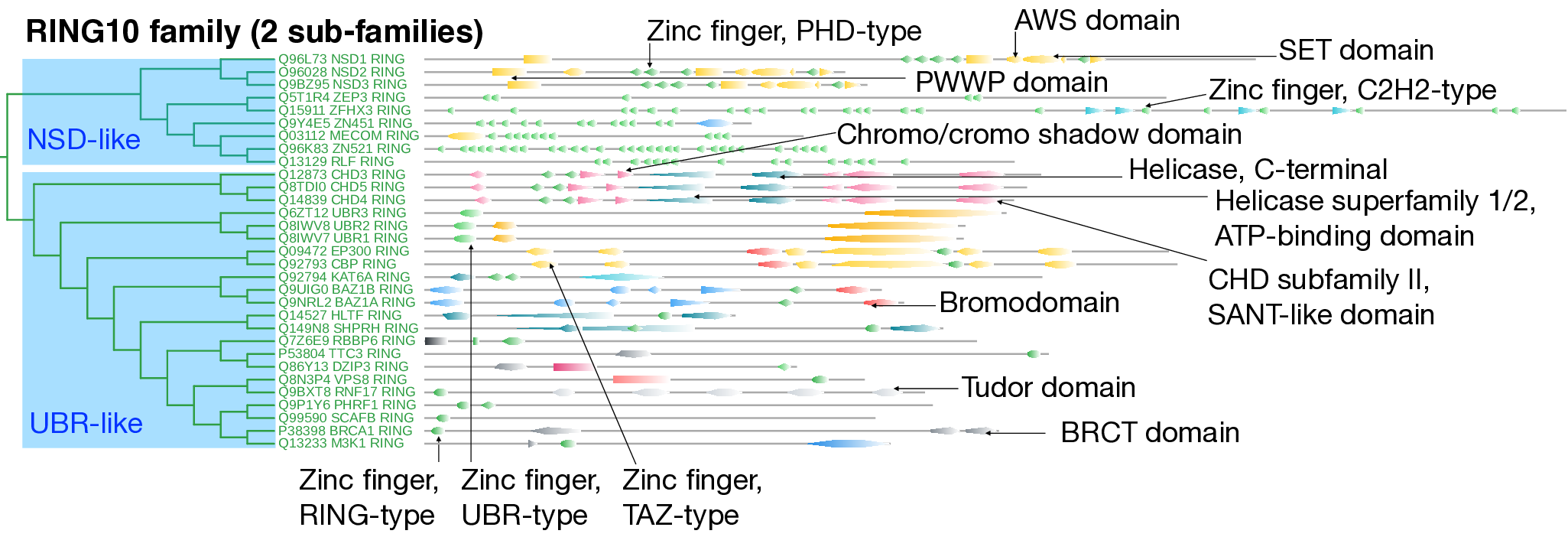
The RING10 family comprises 31 E3 ligases, each exceeding 1000 residues in length. The catalytic domains within this family include RING-, PHD-, UB-R, TAZ-, and C2H2-type Zn-finger domains. Based on domain architecture and sequence similarity, this family is subdivided into two distinct subfamilies.
The first subfamily, "NSD-like," includes NSD1–3 (nuclear receptor-binding SET domain proteins), which are characterized by multiple functional domains such as the PWWP domain, Zn-finger PHD-type domain, SET domain, and NSD CYS-HIS domain. A defining feature of this subfamily is the repetition of Zn-finger PHD-type domains, which are frequently implicated in chromatin interactions. Notably, the SET and PHD-type Zn-finger domains are strongly associated with histone modification and methyltransferase activity, highlighting their potential regulatory role in epigenetic processes.
The second subfamily, "UBR-like," includes CHD, UBR, and BAZ1 ligases, along with other singletons. CHD ligases contain chromodomains, helicase superfamily 1/2 ATP-binding domains, CHD subfamily II-specific features, and SANT-like domains, suggesting roles in chromatin remodeling and gene regulation. The UBR ligases are distinguished by the UBR-like C-terminal domain (IPR044046), forming a prominent subgroup within this family. Ligases such as BAZ1 exhibit a multidomain architecture, including WSTF/Acf1/Cbp146 domains, DDT domains, WHIM1 and WHIM2 domains, PHD-type Zn-finger domains, bromodomains, Tudor domains, and BRCT domains, indicating multifunctional roles in chromatin interactions, DNA repair, and transcriptional regulation. Several members of this subfamily contain the Zn-finger PHD-finger domain, which is commonly found in nuclear proteins that interact with chromatin. The enrichment of various domain types within this subfamily suggests a role in chromatin remodeling, as further supported by the RING10 family's association with nuclear chromosome and histone acetyltransferase complex cellular components, as well as chromatin-binding and histone-binding molecular functions.
However, the heterogeneity of these subfamilies is evident from the multiple sequence alignment (MSA), which exhibits numerous gaps. RING10 members are enriched in biological processes such as DNA metabolism, ubiquitin-dependent protein catabolic processes via the N-end rule pathway, and protein acetylation. They are localized to the SWI/SNF superfamily-type complex, histone acetyltransferase complexes, and the PML body, where they mediate histone-modifying activity, helicase activity, and chromatin binding.
The PDB contains 296 resolved structures covering 21 out of 31 RING10 proteins. Most RING10 family proteins are likely to function as standalone ligases, with only three operating as part of a complex, while the remaining seventeen ligases remain unclassified. Analysis of the ESI network for this family revealed 31 unique E3-specific substrates and four family-specific substrates. Two E3 handles exist for UBR1. Our analysis uncovered new E3s binders targeting NSD2, CHD4, M3K1, CBP, EP300, BAZ1A, KAT6A, NSD3, and BRCA1. Several E3 binders target CBP and EP300, which are co-clustered. Several of these E3 binders are unique and form distinct clusters (KAT6A targeting) and could be developed into lead compounds or repurposed to target related E3s.
For updated information and bibliography, please consult the supplementary information in Dutta et al. (2025, preprint)
The values represent E3 ligase confidence scores, calculated as the fraction of databases that annotate the protein as an E3 ligase (see preprint).